Uciążliwość zapachowa/Węch człowieka/Elementy fizjologii narządu węchu
| « | *** Elementy fizjologii narządu węchu |
» |
| Elementy anatomii narządu węchu | Receptory węchowe i kodowanie zapachu |
Elementy fizjologii narządu węchu
edytujMechanizm percepcji zapachów jest zagadnieniem interdyscyplinarnym, mieszczącym się m.in. w zakresie zainteresowań biofizyków, biochemików, neurobiologów. Na drodze do dzisiejszego stanu wiedzy przełomowe znaczenie miały odkrycia dokonane w XX w., dotyczące budowy i funkcji błony komórkowej. Pierwsze takie modele opisano w pierwszej połowie XX w., np. model dwuwarstwy lipidowej (Gortel i Grendel, 1925) lub błony trójwarstwowej z warstwą białkową (Dowson i Danieli, 1935)[1]. W roku 1972 S. J. Singer i G. L. Nicolson opublikowali model dynamicznej płynnej mozaiki[2][1]. Model był intensywnie rozwijany w następnych latach m.in. przez fizjologów, lekarzy i chemików, których osiągnięcia zostały wyróżnione Nagrodą Nobla, np. za badania:
- dotyczące funkcji pojedynczych kanałów jonowych w komórkach (Erwin Neher; 1991)
- dotyczące fosforylacji białek, jako biologicznego mechanizmu regulatorowego (Bert Sakmann, Edmond H. Fischer, Edwin G. Krebs; 1992)
- białek G i roli tych białek w przekazywaniu sygnału w komórkach (Alfred G. Gilman, Martin Rodbell; 1994)
- dotyczące przekazywania sygnału w układzie nerwowym (Arvid Carlsson, Paul Greengard, Eric R. Kandel; 2000)
- kanałów w błonach komórkowych (Martin Chalfie, Roderick MacKinnon, 2003)
- białka o zielonej fluorescencji (Osamu Shimomura, Martin Chalfie, Roger Y. Tsien, 2008).
Do największego przełomu doprowadziły badania Lindy B. Buck i Richarda Axela wyróżnionych w 2004 roku Nagrodą Nobla za badania receptorów węchowych i organizacji układu węchowego[3].
Dotychczasowe osiągnięcia dotyczyły przede wszystkim pierwszego etapu procesu percepcji zapachu. Zasady działania wyższych pięter analizatora wrażeń węchowych są nadal przedmiotem badań, np. metodami elektroolfaktografii (EOG), neurografii (MRN), psychofizyki, psychofizjologii.
Neuron
edytujAnalizator węchowy człowieka wchodzi w skład układu nerwowego – nerwów czaszkowych. Składa się z komórek nerwowych o różnych kształtach. Niezależnie od kształtu każdy neuron składa się z ciała komórki (perikarion) oraz dendrytów, odbierających bodźce z zewnątrz, i aksonów, odprowadzających impulsy nerwowe w kierunku innych komórek sieci.
W części odpowiadającej za odbieranie sygnałów węchowych z otoczenia ciało neuronów ma tylko dwie wypustki (neurony dwubiegunowe) – jeden dendryt z urzęsioną kolbką przy powierzchni nabłonka oraz jeden akson, tworzący z aksonami innych komórek wiązki przenikające przez kość sitową do jamy czaszki. Zakończenie aksonu jest silnie rozgałęzione (neurony I tworzą z neuronami II liczne synapsy akson-dendryt w kłębuszkach opuszki węchowej).
W częściach analizatora odpowiedzialnych za interpretację sygnałów o odebranych bodźcach chemicznych neurony są silnie rozgałęzione. Zapewnia to równoległe połączenia z różnymi ośrodkami mózgu, odpowiedzialnymi za odczuwanie emocji, rozpoznawanie zapachu oraz nieświadome reakcje organizmu na informacje o chemicznym składzie powietrza otoczenia.
-
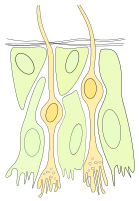 Schemat nabłonka węchowego
Schemat nabłonka węchowego -
 Neuron piramidalny hipokampa
Neuron piramidalny hipokampa -
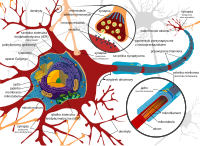 Schemat neuronu i synapsy
Schemat neuronu i synapsy



Budowa błony neuronu
edytujBudowę błony komórkowej ilustruje model „płynnej mozaiki” – dwuwarstwy lipidowej, utworzonej przez ułożone równolegle cząsteczki lipidów. Są to najczęściej fosfolipidy (fosfatydy, fosfotłuszczowce), zbudowane z gliceryny, wyższych kwasów tłuszczowych (nasyconych lub nienasyconych) i kwasu fosforowego związanego z zasadą azotową, np. choliną. Podobne dwuwarstwy powstają spontanicznie w roztworach wodnych, wskutek zróżnicowania sił oddziaływania między cząsteczkami wody i poszczególnymi fragmentami fosfolipidów lub glikolipidów. Hydrofobowe węglowodorowe łańcuchy kwasów tłuszczowych kierują się ku sobie, tworząc wewnętrzną część błony. Hydrofilowe ”głowy” cząsteczek kierują się w kierunku roztworów wodnych (cytoplazma i płyn zewnątrzkomórkowy). Płynną mozaikę tworzą umieszczone w dwuwarstwie lipidowej cząsteczki białek, spełniające różne funkcje fizjologiczne, np. chemoreceptory, kanały jonowe, pompy sodowo-potasowe. Charakterystyczną cechą neuronów jest ich pobudliwość – konsekwencja procesów polaryzacji i depolaryzacji błony komórkowej[4][5].
-

-
 Schemat dwuwarstwy lipidowej
Schemat dwuwarstwy lipidowej -
 Schemat „płynnej mozaiki”
Schemat „płynnej mozaiki”



Polaryzacja i depolaryzacja błony neuronu
edytujCharakterystyczną cechą błony komórkowej neuronów jest ich pobudliwość, czyli zdolność do zmian różnicy potencjałów elektrycznych, występującej między zewnętrzną i wewnętrzną powierzchnią błony. Wartość potencjału błony neuronu niepobudzonego wynosi ok. -60 mV. Jest on utrzymywany na w przybliżeniu stałym poziomie dzięki ustaleniu się szybkości transportu jonów przez błonę – do jej wnętrza i na zewnątrz (równowaga dynamiczna).
W przenoszeniu jonów przez błonę uczestniczą białka:
- tworzące kanały jonowe, umożliwiające prostą dyfuzję z roztworu o stężeniu większym do do roztworu o stężeniu mniejszym,
- uczestniczące w transporcie aktywnym, czyli w przenoszeniu jonów w przeciwnym kierunku, wbrew gradientowi stężeń, np. pompa sodowo-potasowa lub zamykane i otwierane „bramki jonowe” (gates) typu uniport, symport lub antyport.
Z punktu widzenia przenoszenia impulsów nerwowych największe znaczenie mają:
- pompa sodowo-potasowa (Na+/K+ ATPaza) – enzym transportujące jony potasu z cytoplazmy na zewnątrz komórki i jony sodu do wnętrza dzięki energii hydrolizy cząsteczki ATP (transport I rodzaju),
- kanały sodowe i potasowe – cząsteczki białek integralnych, przez które mogą dyfundować jony sodu lub potasu.
W przypadku braku bodźców, gdy utrzymuje się potencjał spoczynkowy spolaryzowanej błony:
- stężenia jonów K+ i Cl- są wielokrotnie większe wewnątrz komórki niż na zewnątrz,
- stężenie Na+ jest wielokrotnie mniejsze wewnątrz niż na zewnątrz.
Struktura obu rodzajów kanałów jonowych zależy od potencjału – jego zmiana (np. pod wpływem bodźca chemicznego lub elektrycznego) może powodować „otworzenie się bramki”, czyli umożliwić wypływanie jonów potasu na zewnątrz i wpływanie jonów sodu do wnętrza komórki.
-
 Zasada działania kanału Na+ bramkowanego acetylocholiną
Zasada działania kanału Na+ bramkowanego acetylocholiną -
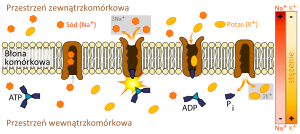 Zasada działania pompy sodowo-potasowej
Zasada działania pompy sodowo-potasowej




Pobudzenie neuronu rozpoczyna się od zmiany konformacji kanału sodowego (otwarcie bramki). Wskutek wpływania jonów sodu do wnętrza zmienia się potencjał elektryczny błony (depolaryzacja), co sprzyja otwieraniu się kolejnych kanałów sodowych. W czasie ok. 1 milisekundy potencjał zmienia się z ok. -60 mV na ok. +40 mV (potencjał czynnościowy). W kolejnym etapie zamykają się kanały sodowe, a otwierają potasowe – wypływanie jonów potasu powoduje powrót do początkowej wartości potencjału w czasie kilku milisekund. Strefa depolaryzacji błony przesuwa się w kierunku zakończenia aksonu, ponieważ lokalna zmiana potencjału wywołuje otwieranie się bramek sodowych w sąsiedniej części spolaryzowanej błony.
Przedstawiony opis wskazuje, że pobudzenie neuronu (powyżej wartości progowej, potrzebnej do wywołania lawinowego wypływu Na+) i przejście fali depolaryzacji wzdłuż neuronu powoduje zmianę stężeń Na+ i K+ w cytoplazmie. Pierwotne wartości są odtwarzane dzięki działaniu pompy Na-K (okres refrakcji)[4][6][5].
Pompa sodowo-potasowa jest białkiem integralnym złożonym z dwóch podjednostek (α i β). Do podjednostki α mogą przyłączać się trzy jony sodowe po wewnętrznej stronie błony. Pod wpływem ATP białko ulega fosforylacji (z utworzeniem ADP), której towarzyszy przemiana konformacyjna i uwolnienie jonów Na+ po stronie zewnętrznej. Utworzenie kompleksu z dwoma jonami potasu powoduje defosforylację i ponowną zmianę konformacji, pozwalającą na uwolnienie jonów K+ po wewnętrznej stronie błony. Jeden cykl przemian trwa ok. 10 milisekund (ok. 100 obrotów/s), co pozwala przetransportować 300 jonów Na+/s i 200 jonów K+/s.
Synapsy
edytujSynapsa jest miejscem, w którym zakończenia aksonów niemal stykają się z błoną komórkową innych neuronów, np. z zakończeniami ich dendrytów. Pomiędzy oboma komórkami znajduje się szczelina synaptyczna. Potencjał czynnościowy, który dociera do zakończenia aksonu pobudzonego neuronu, powoduje egzocytozę pęcherzyków synaptycznych i uwolnienie ich zawartości do szczeliny. Pęcherzyki zawierają związki chemiczne nazywane neuroprzekaźnikami (neurotransmiterami, mediatorami), które dyfundują do błony postsynaptycznej (błony drugiego neuronu) i mogą wywołać jego pobudzenie.
Sieci neuronowe
edytuj
Przekazywanie informacji o odebranym bodźcu kolejnym neuronom odbywa się zgodnie z opisanymi w uproszczeniu zasadami w całym analizatorze węchowym i całym układzie nerwowym, w którym poza opisanymi synapsami akson-dendryt występują liczne . Złożone mechanizmy procesy rozpoznawania zapachów nie zostały dotychczas ostatecznie wyjaśnione. Zarys teorii opracowanej przez Lindę B. Buck i Richarda Axela (Nagroda Nobla z roku 2004 za badania receptorów węchowych i organizacji układu węchowego) opisano w skrócie w rozdziale Receptory węchowe i kodowanie zapachu[3]. Omawianie fizjologicznych podstaw aktywności elektrycznej całego somatycznego układu nerwowego i mózgu wykracza poza ramy niniejszego podręcznika[7].
Elektroolfaktografia
edytujJedna z metod badań narządu węchu nazywana – elektroolfaktografia (EOG, zob. uwaga 1) – polega na rejestracji bioelektrycznej czynności neuronów drogi węchowej – od nabłonka węchowego do mózgu w czasie percepcji zapachu, .
Badania potencjałów błony pojedynczych neuronów
edytujNajwcześniejsze pomiary elektrycznej aktywności pojedynczych neuronów (John Zachary Young 1937) dotyczyły olbrzymich aksonów kałamarnicy.

Ich wymiary pozwalały na umieszczenie elektrod wewnątrz komórki. Wielokrotnie publikowano również wyniki badań potencjałów wywoływanych przez odoranty w komórkach receptorowych owadów z "węchowymi antenami grzebieniastymi". Od połowy XX wieku są publikowane wyniki badań węchu ćmy jedwabnika morwowego, bardzo wrażliwego na zapach bombikolu. Badania tych owadów trwają do dzisiaj. Zapisy zmian potencjałów są nazywane elektroantennogramami (EAG)[9].
Elektroantennogramy były rejestrowane (po odcięciu anten) przy użyciu szklanej elektrody Ag-AgCl z kluczem elektrolitycznym. Równocześnie zapisywano chromatogram powietrza opływającego antenę (zapis GC-EAG).
Badania potencjałów "sumowanych"
edytujŁatwiejsze i częściej stosowane metody badań elektroolfaktograficznych polegają na rejestrowaniu różnicy potencjałów między elektrodą umieszczoną na powierzchni nabłonka węchowego i elektrodą odniesienia[10][11][12].
Elektrody umieszcza się w określonych miejscach nabłonka węchowego zwierząt doświadczalnych (najczęściej szczurów) lub człowieka[13][14]. W czasie otolaryngologicznych badań diagnostycznych stosowano do niedawna wyłącznie metody inwazyjne (w znieczuleniu ogólnym). Obecnie, dzięki udoskonaleniom sprzętu, np. miniaturyzacji sond i endoskopów[15], badania mogą być prowadzone bez znieczulenia. Stwierdzono również, że podobne zapisy EOG można otrzymać umieszczając elektrody pomiarowe na zewnątrz – u nasady nosa (elektroolfaktografia nieinwazyjna)[16][17].
Uniknięcie narkozy umożliwia kontakt z pacjentem w czasie badania – gromadzenie informacji o subiektywnych wrażeniach węchowych odbieranych w czasie zapisu EOG.
Badania OERP (wywołane przez odoranty potencjały EEG)
edytuj
W czasie badań elektroencefalograficznych są rejestrowane zmiany, jakim ulega zapis EEG mózgu pacjenta w czasie wąchania prezentowanych próbek (zapis OERP, od "Olfactory Event-Related Potentials"). W celu zarejestrowania potencjałów wywołanych przez bodźce zapachowe elektrody umieszcza się na skórze głowy w okolicy ciemieniowej i wyspy. Bodźce muszą być powtarzane wielokrotnie, co powoduje że nagrywanie OERPs jest długotrwałe. Rejestrowany jest tzw. efekt Bergera, czyli zahamowania rytmu alfa. Wyniki nie zawsze są jednoznaczne. Zdarza się, że zmiany w EEG nie występują, gdy pacjent odczuwa tylko słaby zapach[12].
Czy znasz odpowiedzi?
edytuj- Jakie elementy tworzą „płynną mozaikę” – błonę komórkową?
| Odpowiedź |
|---|
|
- Jaka energia jest wykorzystywana w czasie aktywnego transportu jonów przez błonę komórkową?
| Odpowiedź |
|---|
- Jaką funkcję pełni w synapsach acetylocholina ?
| Odpowiedź |
|---|
|
- Czym jest impuls nerwowy i jak się przemieszcza?
| Odpowiedź |
|---|
- Co oznaczają skróty EOG i EAG?
| Odpowiedź |
|---|
|
- Jak wykonuje się i czemu służy zapis OERP (Olfactory Event-Related Potentials)?
| Odpowiedź |
|---|
|
Uwagi
edytuj- Symbol EOG jest też stosowany jako skrót określenia "electrooculography (EOG)" lub "elektrookulografia", odnoszącego się do badań narządu wzroku.
Przypisy
- ↑ 1,0 1,1 Uniwersytet Humanistyczno-Przyrodniczy w Siedlcach, Instytut Biologii: Błony komórkowe (pol.). W: Skrypt: Biologia komórki [on-line]. www.ib.uph.edu.pl. [dostęp 2011-06-02].opublikowany w url
- ↑ Singer S.J., Nicolson G.L.. The fluid mosaic model of the structure of cell membranes. „Science”. 175 (23), s. 720–731, luty 1972. DOI: 10.1126/science.175.4023.720. PMID: 4333397 (ang.).
- ↑ 3,0 3,1 Linda B. Buck, Richard Axel. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. „Cell”. 65, 1991 (ang.).
- ↑ 4,0 4,1 Nerw. W: B.D. Hames, N.M. Hooper: Biochemia (wyd. 2). Warszawa: Wydawnictwo Naukowe PWN, 2002, s. 152, 452–456, seria: Krótkie wykłady. ISBN 83-01-13872-6.
- ↑ 5,0 5,1 Marta Kamińska: Czynność bioelektryczna organizmu ludzkiego (pol.). W: Materiały dydaktyczne Zakładu Biofizyki IIM. Politechnika Łódzka [on-line]. www.biofizyka.p.lodz.pl. [dostęp 2012-02-07].opublikowany w url
- ↑ Jarosław Piersa: Elektrofizjologia neuronu (pol.). W: Materiały dydaktyczne Uniwersytetu Toruńskiego [on-line]. www-users.mat.uni.torun.pl, 2008-11-13 Jarosław. [dostęp 2012-02-09].opublikowany w url
- ↑ Andrzej Wróbel. Zbiorcza aktywność elektryczna mózgu. „Kosmos”. 46 (3), s. 317 326, 1997 (pol.).
- ↑ Antenna (insects). [dostęp 2010-09-16].
- ↑ J. Ziesmann, I. Valterova, K. Haberkorn, M.G. de Brito Sanchez i inni. Chemicals in Laboratory Room Air Stimulate Olfactory Neurons of Female Bombyx mori. „Chemical Senses”. 25. s. 31-37.
- ↑ A. Holley. Przegląd prasy: Recherche VII-VII 1975. „Problemy”. 4, s. 57-59, 1976 (pol.).
- ↑ [2.3. Powstawanie i przewodzenie impulsów nerwowych]. W: J. Kośmider: Dezodoryzacja gazów i ścieków.. Cz. IV.: Oceny zapachowej jakości powietrza i skuteczności dezodoryzacji. Szczecin: Wydawnictwo Naukowe Politechniki Szczecińskiej, 1990, s. 16-17. (pol.)
- ↑ 12,0 12,1 Karolina Dżaman. Współczesne metody badania węchu i smaku. „Otolaryngologia”. 7 (4), s. 173-177, 2008.
- ↑ J.S. Kauer, D.G. Moulton. Responses of olfactory bulb neurons to odor stimulation of small nasal areas in the salamander. „J Physiol”. 243, s. 717–737, 1974.
- ↑ John W. Scott, Tracy Brierley, Frederick H. Schmidt. Chemical Determinants of the Rat Electro-Olfactogram. „The Journal of Neuroscience”. 20 (12), s. 4721-4731, June 2000 (ang.).
- ↑ Izrael:_endoskop_w_postaci_minirobota (pol.). [dostęp 2010-09-16].
- ↑ L. Wang, C. Hari, L. Chen, T. Jacob. A new non-invasive method for recording the electro-olfactogram using external electrodes. „Clin Neurophysiol”. 115, s. 1631–1640, 2004 (ang.).
- ↑ Hadas Lapid, Han-Seok Seo, Benno Schuster, Elad Schneidman i inni. Odorant Concentration Dependence in Electroolfactograms Recorded From the Human Olfactory Epithelium. „J. Neurophysiol”, August 5 2009. American Physiological Society. DOI: 10.1152/jn.91321.2008 (ang.).